乳酸不再是代谢废物,而是能量来源、信号分子和免疫调节因子。乳酸浓度升高常与癌症、炎症和自身免疫病有关。赖氨酸乳酰化(lysine lactylation)这一新兴蛋白质翻译后修饰(PTM)的分子机制及其在癌症、免疫应答、代谢调控等生理病理过程中的作用,特别强调了两类乳酰转移酶(lactyltransferases):
- 乳酰-CoA依赖途径(lactyl-CoA-dependent):如p300、GCN5、KAT8等乙酰转移酶。
- 乳酰-CoA非依赖途径(lactyl-CoA-independent):以AARS1/2(alanyl-tRNA synthetases)为代表的新型乳酸感受器和乳酰转移酶。
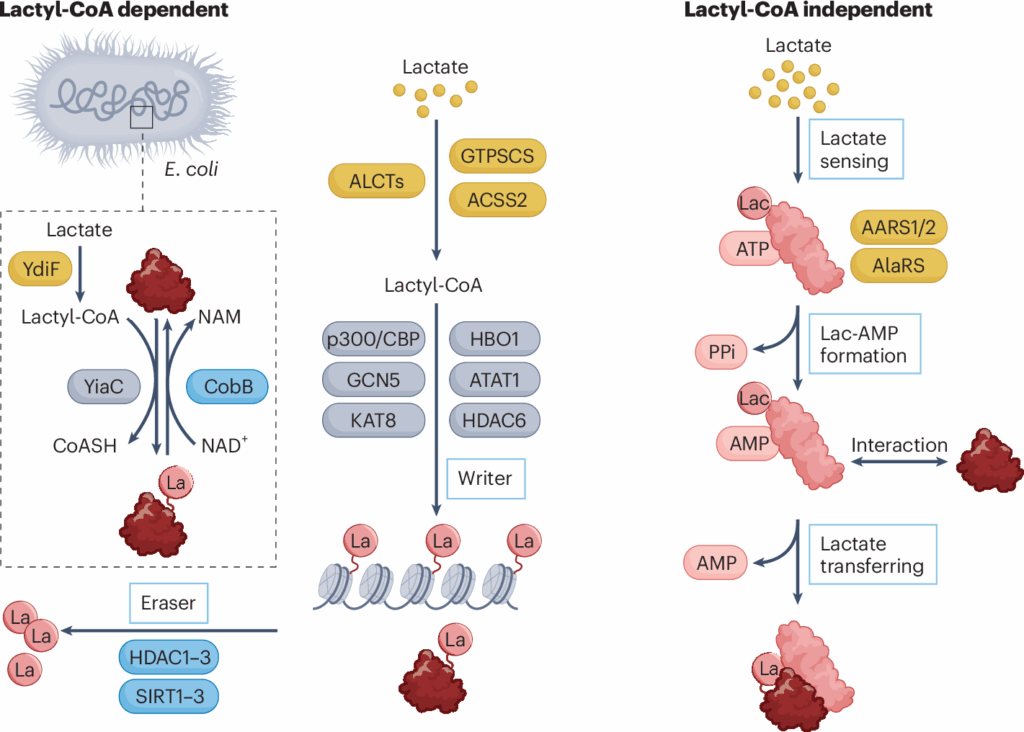
赖氨酸乳酰化(lactylation)的发现与机制
乳酰化是一种新型赖氨酸修饰方式,最初认为通过乳酰-CoA中介。后发现AARS1/2可直接催化乳酸+ATP生成Lac-AMP,并将乳酰基转移到蛋白赖氨酸残基上,无需乳酰-CoA。
乳酰化靶标与生物学功能
组蛋白乳酰化(Histone lactylation):如H3K18la、H3K9la、H4K12la——促进转录激活、癌症进展、免疫调节等。
参与疾病:黑色素瘤、结直肠癌、膀胱癌、肝纤维化、心梗、近视、阿尔兹海默病等。
非组蛋白乳酰化(Non-histone lactylation):
修饰蛋白如:YAP、TEAD1、NBS1、PDHA1、CPT2、p53 等,调控细胞周期、DNA修复、氧化磷酸化、癌症进展、化疗耐药等。
乳酰转移酶分类:
乳酰-CoA依赖:p300、CBP、GCN5、KAT8、HBO1、ATAT1等酶均可介导组蛋白乳酰化。然而乳酰-CoA浓度极低,限制其活性。
乳酰-CoA非依赖(新发现):AARS1/2识别乳酸,ATP辅助生成Lac-AMP,中间产物直接用于乳酰化。影响p53、YAP–TEAD1复合体、cGAS等蛋白功能。
去乳酰化酶(Delactylases):HDAC1–3、SIRT1–3具备不同程度的乳酰去除活性。HDAC3活性最高,尤其针对H4K5la。去乳酰化调控能影响癌症免疫、代谢和DNA修复。
治疗潜力与干预策略
| 靶点类别 | 干预方式 | 示例药物或方法 |
|---|---|---|
| LDHA | 抑制乳酸产生 | Oxamate、GSK2837808A |
| MCT1/4 | 抑制乳酸跨膜转运 | AZD3965、VB124 |
| p300、GCN5 等转移酶 | 抑制乳酰化酶活性 | C646、A-485 |
| AARS1/2 | 抑制乳酸结合活性 | β-丙氨酸(β-Ala)、L-丙氨酸(L-Ala) |
| Delactylases | 激活去乳酰化 | Honokiol(SIRT3激动剂) |
这些策略对癌症、免疫病、心血管疾病和代谢性疾病均具有潜在价值。酶选择性机制尚不明确,需进一步探索其底物识别、组织特异性和调控机制。结构生物学研究不足,对催化效率、结构基础了解不全。蛋白功能调控机制待完善,特别是在细胞定位和液-液相分离调控中的作用。
本站原创,如若转载,请注明出处:https://www.ouq.net/3732.html
 微信打赏,为服务器增加50M流量
微信打赏,为服务器增加50M流量  支付宝打赏,为服务器增加50M流量
支付宝打赏,为服务器增加50M流量