
线粒体不仅合成细胞功能所需的能量,还参与多种细胞途径,包括细胞凋亡、钙稳态、炎症和免疫。线粒体是经历裂变和融合循环的动态细胞器,这些碎片化和超融合网络之间的转变确保了线粒体功能,使其能够适应代谢变化或细胞应激。线粒体形态缺陷与多种疾病有关,这凸显了阐明调节线粒体形态的分子机制的重要性。在这里,我们讨论了对核心线粒体动力学蛋白的组装和作用机制的最新结构见解,例如控制分裂的动力蛋白相关蛋白 1 (DRP1),以及驱动膜融合的线粒体融合蛋白 (MFN1 和 MFN2) 和视神经萎缩 1 (OPA1)。此外,我们还提供了线粒体膜融合和裂变过程中不同蛋白质、脂质和细胞器之间复杂相互作用的最新观点。总体而言,我们的目标是提出一个有价值的框架,反映当前关于线粒体膜重塑如何调节的观点。
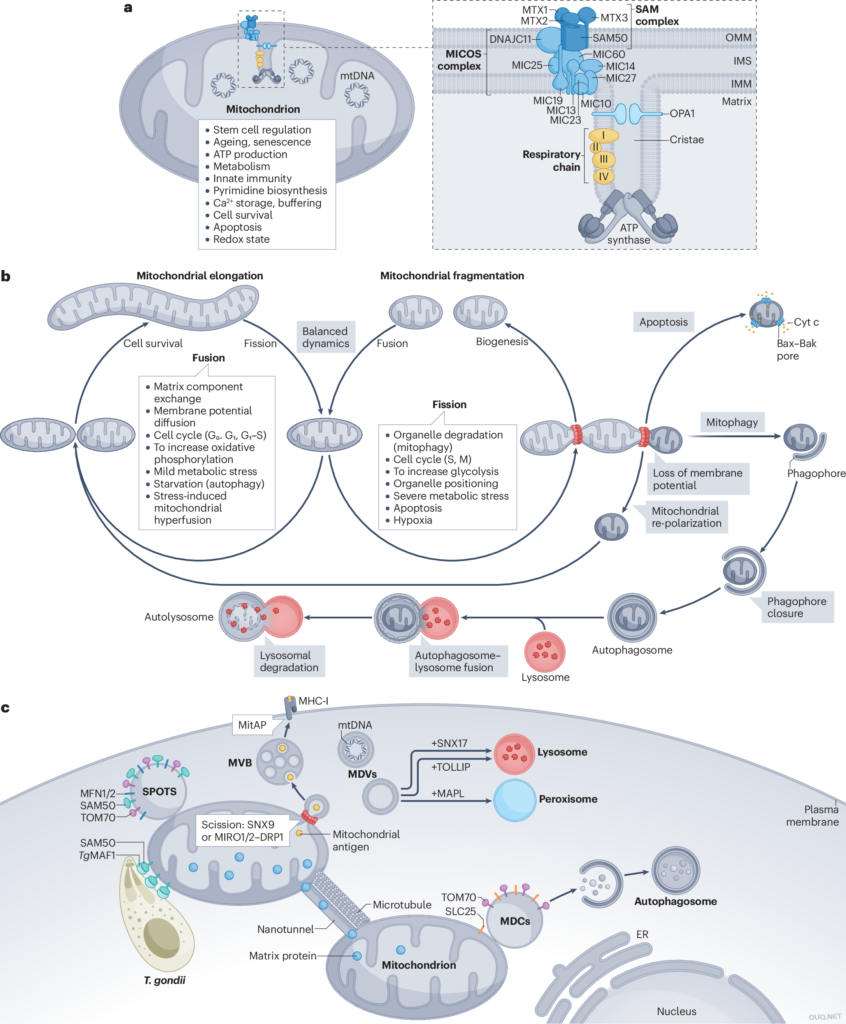
a、线粒体不仅仅是细胞的动力源,还控制着细胞命运的决定,例如干细胞命运、细胞凋亡和细胞存活。图中描绘了线粒体超微结构和生理功能的关键方面。线粒体基质含有线粒体 DNA (mtDNA),被两层膜包围,即线粒体外膜 (OMM) 和线粒体内膜 (IMM),它们被膜间隙 (IMS) 隔开。线粒体接触位点和嵴组织系统 (MICOS) 由亚基 MIC10、MIC13、MIC14、MIC19、MIC25、MIC26、MIC27 和 MIC60 组成,是线粒体膜间隙桥接 (MIB) 复合体的一部分,该复合体还包括分选和组装机制组件 50 同源物 (SAM50)、DNAJC11 和 metaxins1–3 (MTX1–MTX3)。MIB 复合物与动力蛋白样 GTPase 视神经萎缩 1 (OPA1) 和 ATP 合酶二聚体一起控制嵴结构并将 OMM 物理地连接到 IMM。b、在整个生命周期中,线粒体不断经历裂变和融合循环,以应对不同的压力并适应细胞的代谢需求。线粒体伸长与呼吸速率增加有关,被认为是一种保护机制,可保护功能性线粒体免受细胞死亡和轻微应激的影响。线粒体碎裂发生在有丝分裂 (M) 期间,以确保有足够数量的线粒体分配给子细胞,并承受各种严重应激。潜在的应激包括细胞凋亡,这是一种程序性细胞死亡途径,由细胞凋亡调节剂 BAX–Bcl-2 拮抗剂杀伤剂 (BAK) 在 OMM 处的寡聚化诱导,从而驱动细胞色素 c (Cyt c) 释放到细胞溶胶中,或线粒体自噬,涉及自噬体内小的功能障碍线粒体的降解。c,几种不同的过程调节线粒体衍生结构的生物发生和运输。线粒体衍生囊泡 (MDV) 被认为是由特定蛋白质标记的小结构,可能含有或不含有 mtDNA。不同类型的 MDV 靶向多种细胞器,包括过氧化物酶体和溶酶体。MDV 还通过呈递到主要组织相容性复合体 I 类 (MHC-I) 介导线粒体抗原呈递 (MitAP),线粒体抗原通过多泡体 (MVB) 运输到 MHC-I。MDV 的分裂需要内吞相关蛋白分选 nexin-9 (SNX9) 或由线粒体 Rho GTPase 1 (MIRO1) 依赖性或 MIRO2(下文中为 MIRO1/2)依赖性膜管化引发的裂变事件,随后是动力蛋白相关蛋白 1 (DRP1) 驱动的裂变事件。线粒体衍生区室 (MDC) 通过自噬调节选择性 OMM 蛋白(例如,线粒体输入受体亚基 Tom70)和溶质载体家族 25 (SLC25) 成员的循环。弓形虫感染会触发一种未知功能的 OMM 结构的形成,称为线粒体外膜阳性结构 (SPOT)。SPOT 的形成需要弓形虫线粒体相关因子 1 (TgMAF1) 与宿主线粒体输入受体 TOM70 和 OMM 转位酶 SAM50 相互作用。促融合因子线粒体融合蛋白 (MFN) 被隔离在 SPOT 中。纳米隧道以微管依赖的方式在不同的线粒体之间运输基质和膜蛋白。
本文来自,本文观点不代表OUQ技术分享立场,转载请联系原作者。
 微信打赏,为服务器增加50M流量
微信打赏,为服务器增加50M流量  支付宝打赏,为服务器增加50M流量
支付宝打赏,为服务器增加50M流量 



